На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Клеточная стенка или плазматическая мембрана
Бесцветная, прозрачная и очень прочная
Пропускает в клетку и выпускает из клетки вещества.
Клеточная мембрана полупроницаемая
В ней располагаются все другие части клетки
Находится в постоянном движении
Обеспечивает передачу наследственных свойств дочерним клеткам при делении
Сферической или неправильной формы
Принимает участие в синтезе белка
Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок
Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке.
По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль
Используют световую энергию солнца и создают органические из неорганических
Форма дисков, отграниченных от цитоплазмы двойной мембраной
Образуются в результате накопления каротиноидов
Жёлтые, оранжевые или бурые
Содержатся в корнях, клубнях, луковицах
Состоит из двух мембран (наружная и внутренняя) с порами
Отграничивает ядро от цитоплазмы
Даёт возможность осуществляться обмену между ядром и цитоплазмой
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.
Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.
Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
источник
Аппарат Гольджи – одномембранная, микроскопическая органелла эукариотической клетки, которая предназначена для завершения процессов синтеза клетки и обеспечивает вывод образовавшихся веществ.
Исследование структурных компонентов комплекса Гольжи началось еще в 1898 итальянским ученым-гистологом Камилло Гольджи, в честь него органелла и была названа. Изучение органоида проходило впервые в составе нервной клетки.
В пластинчатом комплексе (аппарат Гольджи) имеется три части:
- Цис-цистерна — находится вблизи ядра, постоянно взаимодействует с гранулярной эндоплазматической сетью;
- медиал-цистерна или промежуточная часть;
- транс-цистерна — отдаленная от ядра, дает трубчатые разветвления, формируя транс-сеть Гольджи.
Пластинчатый комплекс в клетках разной природы и даже на различных этапах дифференцировки одной клетки, иногда имеет отличительные черты в строении.
 Строение аппарата Гольджи
Строение аппарата Гольджи
Имеет вид стопки, которая состоит от трех до восьми цистерн, толщиной около 25 нм, они уплощены в центральной части и расширяются в направлении к периферии, напоминают стопку перевернутых тарелок. Поверхности цистерн примыкают друг к другу очень плотно. От периферической части отпочковываются небольшие мембранные пузырьки.
Клетки человека имеют одну, реже пару стопок, а клетки растений могут содержать несколько таких образований. Совокупность цистерн (одна стопка) совместно с окружающими ее пузырьками называется диктиосомой. Несколько диктиосом могут связываться между собой, формируя сеть.
Полярность – наличие цис-стороны, направленной к ЭПС и ядру, где происходит слияние везикул, и транс-стороны, устремленной к клеточной оболочке (это особенность хорошо прослеживается в клетках секретирующих органов).
Асимметричность – сторона расположенная ближе к ядру клетки (проксимальный полюс) вмещает «незрелые» белки, к ней постоянно присоединяются везикулы, отсоединившиеся от ЭПС, транс-сторона (дистальный, зрелый полюс) содержит уже модифицированные белки.
При разрушении чужеродными агентами пластинчатого комплекса, происходит разделение аппарата Гольджи на отдельные части, но его основные функции при этом сохраняются. После возобновления системы микротрубочек, которые были хаотично разбросаны в цитоплазме, части аппарата собираются, и снова превращаются в нормально функционирующий пластинчатый комплекс. Физиологическое разделение происходит и в обычных условиях жизнедеятельности клеток, во время непрямого деления.
ЭПС – это часть комплекса Гольджи?
Однозначно нет. Эндоплазматическая сеть – это самостоятельная мембранная органелла, которая построена из системы замкнутых канальцев, цистерн, сформированных непрерывной мембраной. Основная функция – синтез белков, с помощью рибосом, размещенных на поверхности гранулярной ЭПС.
Существует ряд сходных признаков между ЭПС и аппаратом Гольджи:
- Это внутриклеточные образования, отграниченные от цитоплазмы мембраной;
- отделяют мембранные пузырьки, которые наполнены органическими продуктами синтеза;
- вместе формируют единую синтезирующую систему;
- в секретирующих клетках имеют наибольшие размеры и высокий уровень развития.
Чем образованы стенки эндоплазматической сети и комплекса Гольджи?
Стенки ЭПС и аппарата Гольджи представлены в виде однослойной мембраны. Эти органеллы вместе с лизосомами, пероксисомами и митохондриями объединены в группу мембранных органоидов.
Что происходит в комплексе Гольджи с гормонами и ферментами?
За синтез гормонов отвечает эндоплазматическая сеть, на поверхности ее мембраны идет производство гормональных веществ. В комплекс Гольджи поступают синтезированные гормоны, здесь они накапливаются, затем идет переработка и выведение их наружу. Поэтому в клетках эндокринных органов встречаются комплексы больших размеров (до 10 мкм).
Протеолиз белковых веществ, что приводит к активации белков, так проинсулин переходит в инсулин.
Обеспечивает транспорт из клетки продуктов синтеза ЭПС.
Самой важной функцией комплекса Гольджи считают выведение из клетки продуктов синтеза, поэтому его еще называют транспортным аппаратом клетки.
Синтез полисахаридов, таких как пектин, гемицеллюлоза, которые входят в состав мембран растительных клеток, образование гликозаминогликанов, одного из составляющих межклеточной жидкости.
В цистернах пластинчатого комплекса идет созревание белковых веществ, необходимых для секреции, трансмембранных протеинов клеточной мембраны, ферментов лизосом и др. В процессе созревания белки постепенно перемещаются по отделам органоида, в которых завершается их формирование и происходит гликозилирование и фосфорилирование.
Формирование липоптротеидных веществ. Синтез и накопление слизистых веществ (муцина). Образование гликолипидов, которые входят в состав мембранного гликокаликса.
Передает белки в трех направлениях: к лизосомам (перенос контролируется ферментом – маннозой- 6-фосфат), к мембранам или внутриклеточной среде, и к межклеточному пространству.
Вместе с зернистой ЭПС образует лизосомы, путем слияния отпочковавшихся везикул с автолитическими ферментами.
Экзоцитозный перенос – везикула, подойдя к мембране, встраивается в нее и оставляет свое содержимое с наружной стороны клетки.
источник
или «желудки» клетки
Какую функцию выполняет желудок человека? Пищу переваривает! Так вот у каждой эукариотической клетки есть такой желудочек, и не один!
- В желудке человека кислая среда, и в лизосомах кислая среда, у человека в пищеварении участвуют ферменты, и у лизосом обязательный компонент — ферменты.


- Форма тельца у лизосом разная, но, в основном, это сферические или шарообразные одномембранные органеллы.
- Количество их в клетке тоже разное — от одной (вакуоль у клетки растений и грибов) до нескольких тысяч.
В животных клетках их обычно от нескольких сотен до нескольких тысяч.
Функции лизосом
- Ферментативная обработка поступающих в клетку веществ — расщепление крупных органических молекул на более простые составляющие;
- Это оганеллы, которые могут пожертвовать собой ради спасения клетки! Если в клетку поступил какой-либо яд, лизосома поглощает его фагоцитозом , переваривает, т.е. нейтрализует яд, при этом сама погибает…
На рисунках лизосомы изображают обычно вот так:
Обычно лизосомы образуются из аппарата Гольджи — мембранного компонента клетки.
Аппарат похож на стопку мешочков и систему пузырьков. Со временем эти пузырьки «отпочковываются» — образуются лизосомы.
Выглядит аппарат Гольджи вот так:
И лизосомы, и аппарат Гольджи — компоненты именно эукариотических клеток. Ни у вирусов, ни у бактерий их нет
- Производство лизосом;
- Накопление белков;
- Формирует углеводные компоненты, входящие в состав мембраны клетки;
- Аппарат Гольдджи содержит ферменты;
- Транспорт веществ из Эндоплазматического ретикулума — в аппарате Гольджи белки «специализируются» — белки для мембраны, белки для лизосом и т.д. «Специализация» идет за счет ферментов.
Основные функции Аппарата Гольджи — образование лизосом, сократительных и пищеварительных вакуолей простейших эукариотических организмов, а также формирование компонентов клеточной стенки растений.
Особенно хорошо этот органоид развит в секреторных клетках, например, в вырабатывающих слизь клетках кишечника.
источник
Аппарат Гольджи, также называемый комплексом Гольджи — органелла встречающаяся, как в клетках растений, так и животных, и обычно состоит из совокупности чашеобразных отделов с мембраной, называемых цистернами, которые выглядят как стопка сдутых воздушных шаров.
Однако у некоторых одноклеточных жгутиковых имеется 60 цистерн, формирующих аппарат Гольджи. Точно так же количество стопок комплекса Гольджи в клетке изменяется в зависимости от ее функций. Клетки животных, как правило, содержат от 10 до 20 стопок на одну клетку, объединенных в один комплекс трубчатыми соединениями между цистернами. Аппарат Гольджи обычно расположен близко к ядру клетки.
Из-за относительно больших размеров комплекс Гольджи был одной из первых наблюдаемых органелл в клетках. В 1897 году итальянский врач по имени Камилло Гольджи, изучающий нервную систему, использовал новую технологию окрашивания, которую сам же разработал (и которая актуальна в наши дни). Благодаря новому методу, ученый смог разглядеть клеточную структуру и назвал ее внутренним ретикулярным аппаратом.
Вскоре после того, как он публично объявил о своем открытии в 1898 году, структура была названа в его честь, становясь универсально известной как аппарат Гольджи. Тем не менее, многие ученые того времени не верили, что Гольджи наблюдал настоящую органеллу клетки, и списывали открытие ученного на визуальное искажение, вызванное окрашиванием. Изобретение электронного микроскопа в двадцатом веке окончательно подтвердило, что аппарат Гольджи является клеточной органеллой.
У большинства эукариот аппарат Гольджи формируется из стопок мешочков, состоящих из двух основных отделов: цис-отдела и транс-отдела. Цис-отдел представляет собой комплекс сплюснутых мембранных дисков, известных как цистерны, происходящие из везикулярных кластеров, которые устремляются из эндоплазматического ретикулума.
Клетки млекопитающих обычно содержат от 40 до 100 стопок. Как правило, от в каждая стопка включает от 4 до 8 цистерн. Однако у некоторых протистов наблюдается около 60 цистерн. Этот набор цистерн разбит на цис, медиальные и транс-отделы. Транс-отдел представляет собой конечную цистернальную структуру, из которой белки упаковываются в везикулы, предназначенные для лизосом, секреторных везикул или клеточной поверхности.
Аппарат Гольджи часто считается отделом распределения и доставки химических веществ клетки. Он модифицирует белки и липиды (жиры), которые продуцируются в эндоплазматическом ретикулуме, и готовит их для экспорта за пределы клетки или для транспортировки в другие места внутри клетки. Белки и липиды, построенные в гладком и шероховатом эндоплазматическом ретикулуме, укладываются в крошечные пузырьковые везикулы, которые движутся через цитоплазму, пока не достигнут комплекса Гольджи.
Везикулы сливаются с мембранами Гольджи и высвобождают, содержащиеся внутри молекулы в органеллу. Оказавшись внутри, соединения дополнительно обрабатываются с помощью аппарата Гольджи, а затем направляются в везикуле к месту назначения внутри или вне клетки. Экспортируемые продукты представляют собой секреции белков или гликопротеинов, которые являются частью функции клетки в организме. Другие вещества возвращаются в эндоплазматический ретикулум или могут созревать, чтобы впоследствии стать лизосомами.
Модификации молекул, которые осуществляются в комплексе Гольджи, происходят упорядоченным образом. Каждая цистерна имеет два основных отдела: цис-отдел — это конец органеллы, где вещества поступают из эндоплазматического ретикулума для обработки, а также транс-отдел, где они выходят в форме меньших отдельных везикул. Следовательно, цис-отдел расположен вблизи эндоплазматического ретикулума, откуда поступает большая часть веществ, а транс-отдел расположен вблизи плазматической мембраны клетки, куда отправляются многие из веществ, модифицирующиеся в аппарате Гольджи.
Химический состав каждого отдела, а также ферменты, содержащиеся в люменах (внутренних открытых пространствах цистерн) между отделами, являются отличительными. Белки, углеводы, фосфолипиды и другие молекулы, образующиеся в эндоплазматическом ретикулуме, переносятся на аппарат Гольджи, чтобы подвергнутся биохимическому модифицированию при переходе от цис к транс-отделам комплекса. Ферменты, присутствующие в люмене Гольджи, модифицируют углеводную часть гликопротеинов путем добавления или вычитания отдельных мономеров сахара. Кроме того, аппарат Гольджи сам по себе производит самые разнообразные макромолекулы, включая полисахариды.
Комплекс Гольджи в растительных клетках продуцирует пектины и другие полисахариды, необходимые для структуры растений и обмена веществ. Продукты, экспортируемые аппаратом Гольджи через транс-отдел, в конечном итоге сливаются с плазматической мембраной клетки. Среди наиболее важных функций комплекса — сортировка большого количества макромолекул, продуцируемых клеткой, и их транспортировка в необходимые пункты назначения. Специализированные молекулярные идентификационные метки или метки, такие как фосфатные группы, добавляются ферментами Гольджи, чтобы помочь в этом процессе сортировки.
источник
Аппарат Гольджи, он же комплекс Гольджи, представляет собой один из важнейших компонентов в строении клетки. Эта клеточная органелла, названая в честь итальянского биолога Камилло Гольджи, который ее открыл в 1898 году, она имеет вид комплекса полостей, ограниченных одиночными мембранами. По сути, аппарат Гольджи это мембранная структура эукариотической клетки.
Если мы посмотрим на аппарат Гольджи в электронный микроскоп, то увидим него нечто напоминающее стопку наложенных друг на друга мешочков, около которых находится множество пузырьков. В середине каждого подобного мешка находится узкий канал, который расширяется на концах в так званые цистерны. От них в свою очередь отпочковываются пузырьки. Вокруг центральной стопки образуется система связанных между собой трубочек.
Внешняя сторона аппарат Гольджи имеет немного выпуклую форму, там наши стопки образуют новые цистерны путем слияния пузырьков отпочковывающихся от гладкой эндоплазматической сети. С внутренней стороны аппарата цистерны завершают свое созревание и также распадаются вновь на пузырьки. Подобным образом происходит перемещение цистерн (мешочков, стопок) от наружной стороны органеллы к внутренней.
Также часть комплекса Гольджи, которая располагается ближе к ядру клетки, называется «цис», а часть, которая находится ближе к мембране, называется «транс».
Так выглядит аппарат Гольджи на рисунке.
Роль аппарата Гольджи в жизни клетки разнообразна, в основном она сводится к модификации и перераспределению синтезирующих веществ и также их выведению за пределы клетки, образованию лизосом и построению цитоплазматической мембраны.
Весьма высока активность аппарата Гольджи в секреторных клетках. Белки, которые поступающие из эндоплазматической сети концентрируются в аппарате Гольджи, затем в пузырьках Гольджи переносятся к мембране.
В клетках растений при формировании клеточной стенки именно Гольджи секретирует углеводы, которые служат матриксом для нее. При помощи микротрубочек отпочковавшиеся пузырьки Гольджи перемещаются и их мембраны сливаются с цитоплазматической мембраной, а содержимое включается в клеточную стенку.
Комплекс Гольджи бокаловидных клеток (они находятся в толще эпителия слизистой оболочки кишечника и дыхательных путей) секретирует гликопротеин муцин, он образует слизь.
А в клетках кишечника именно аппарат Гольджи выполняет важную функцию по перемещению липидов. Происходит это таким образом: жирные кислоты и глицерол попадают в клетки, затем в эндоплазматической сети происходит синтез своих липидов, большая часть их которых покрывается белками и при помощи Гольджи транспортируется к клеточной мембране, пройдя через которую липиды окажутся в лимфе.
Также благодаря аппарату Гольджи происходит формирование лизосом, на которых более детально остановимся в будущей статье.
И в завершение образовательное видео по теме нашей статьи.
источник
Аппарат Гольджи – система мембран, напоминающих вогнутые стопки. Каждая стопка – своеобразная цистерна, мешочек, полость, образованная слиянием двух мембран. Это структурная единица органоида, которая называется диктиосомой. В одной органелле число диктиосом может варьировать от четырёх до семи.
Рис. 1. Сроение комплекса Гольджи.
Цистерны взаимодействуют между собой посредством системы трубочек и пузырьков. По структуре и функциональному назначению аппарат Гольджи делится на три отдела. В каждом отделе находятся определённые ферменты, которые участвуют в модификации, попавших в органеллу веществ. Процесс начинается с цис-отдела. Краткое описание каждого отдела представлено в таблице “Строение и функции комплекса Гольджи в клетке”.
Происходящие процессы
Цис-отдел, расположенный около ядра
Осуществляется фосфорилирование белков
Медиальный или срединный состоит из нескольких цистерн
Сортировка и модификация веществ
Транс-отдел – конечный отдел, удалённый от ядра и близко расположенный к клеточной мембране – плазмалемме
Синтез полисахаридов, образование трёх групп белков – секреторных, регенерационных, лизосомальных, выход сформировавшихся пузырьков
В животной клетке комплекс Гольджи расположен ближе к ядру и часто соприкасается с шероховатой эндоплазматической сетью (ЭПС). В растительных клетках цистерны рассеяны по цитоплазме.
Органоид выполняет три важных функции:
- перенос и преобразование белков;
- формирование и модификация полисахаридов и липидов;
- производство лизосом.
Работа комплекса Гольджи не до конца понятна биологам. Главная функция органеллы – синтез секретов, которые в дальнейшем транспортируются наружу. Большинство секретов имеют белковое происхождение, поэтому комплекс Гольджи перерабатывает первичные, незрелые белки, отделившиеся от ЭПС, в готовые секреты. Механизм этого преображения и особенности процесс транспортировки белков через все отделы до конца не ясны.
Аппарат Гольджи производит гликолипиды – сложные соединения, образованные углеводами и жирами. Основу веществ составляют полисахариды, к которым прикрепляются остатки жирных кислот. Гликолипиды входят в состав нервных тканей и клеточных мембран.
Третья важная функция – производство лизосом. Они также «изготовляются» из белков ЭПС. Аппарат Гольджи формирует первичные лизосомы – органеллы, напоминающие пузырёк или везикулу. Снаружи лизосома ограничена тонкой мембраной, внутри находятся ферменты, расщепляющие органические вещества, которые поступают снаружи или производятся клеткой (продукты жизнедеятельности). Отделившиеся от комплекса Гольджи первичные лизосомы сливаются в цитоплазме с твёрдыми или жидкими веществами, превращаясь во вторичные лизосомы, которые выполняют функцию переваривания.
Рис. 3. Процесс образования лизосом.
Комплекс Гольджи наиболее развит в клетках, выделяющих различные секреты.
Аппарат Гольджи – важная органелла растительных и животных клеток. Она состоит из мембран, образующих полости и сложенных стопкой. Через полости комплекса Гольджи проходят белки, жиры, липиды, из которых образуются сложные соединения, участвующие в жизнедеятельности клетки и организма в целом. Аппарат Гольджи производит «строительный» материал из углеводов и липидов, секреты, ферменты, лизосомы.
источник
Комплекс, или аппарат, Гольджи назван так в честь открывшего его ученого. Это клеточная органелла имеет вид комплекса полостей, ограниченных одинарными мембранами. В растительных клетках и у простейших представлен несколькими отдельными более мелкими стопками (диктиосомами).
Комплекс Гольджи по внешнему виду, видимому в электронный микроскоп, напоминает стопку наложенных друг на друга дискообразных мешочков, около которых находится множество пузырьков. Внутри каждого «мешка» находится узкий канал, расширяющийся на концах в так называемые цистерны (иногда цистерной называют весь мешочек). От них отпочковываются пузырьки. Вокруг центральной стопки формируется система взаимосвязанных трубочек.
С наружней, имеющей несколько выпуклую форму, стороны стопки образуются новые цистерны путем слияния пузырьков отпочковывающихся от гладкой эндоплазматической сети. На внутренней стороне цистерны завершают свое созревание и распадаются снова на пузырьки. Таким образом, цистерны (мешочки стопки) Гольджи перемещаются от наружней стороны к внутренней.
Часть комплекса, располагающаяся ближе к ядру, называется «цис». Та, что ближе к мембране, – «транс».
Функции аппарат Гольджи разнообразны, в общей сложности сводятся к модификации, перераспределению синтезированных в клетке веществ, а также их выведению за пределы клетки, образованию лизосом и построению цитоплазматической мембраны.
Активность комплекса Гольджи высока в секреторных клетках. Белки, поступающие из ЭПС, концентрируются в аппарате Гольджи, затем переносятся к мембране в пузырьках Гольджи. Ферменты секретируются из клетки путем обратного пиноцитоза.
К приходящим в Гольджи белкам присоединены олигосахаридные цепочки. В аппарате они модифицируются и служат маркерами, с помощью которых белки сортируются и направляются по своему пути.
У растений при формировании клеточной стенки Гольджи секретирует углеводы, служащие матриксом для нее (целлюлоза здесь не синтезируется). Отпочковавшиеся пузырьки Гольджи перемещаются с помощью микротрубочек. Их мембраны сливаются с цитоплазматической мембраной, а содержимое включается в клеточную стенку.
Комплекс Гольджи бокаловидных клеток (находятся в толще эпителия слизистой оболочки кишечника и дыхательных путей) секретирует гликопротеин муцин, который в растворах образует слизь. Подобные вещества синтезируются клетками кончика корня, листьев и др.
В клетках тонкого кишечника аппарат Гольджи выполняет функцию транспорта липидов. В клетки попадают жирные кислоты и глицерол. В гладкой ЭПС происходит синтез своих липидов. Большинство из них покрываются белками и посредством Гольджи транспортируются к клеточной мембране. Пройдя через нее, липиды оказываются в лимфе.
Важной функцией является формирование лизосом.
источник
Лабораторно-практическое занятие № 9
Тема: «Аппарат (комплекс) Гольджи»
Цель занятия: выявить морфо-функциональные особенности комплекса Гольджи.
1. Тонкое строение аппарата Гольджи.
1. Фотографии, схемы, рисунки из Атласа по биологии клетки, Ж.-К.Ролана, А. Селоши, Д. Селоши, пер.В.П. Белого, под ред. Ю.С. Ченцова. ─ М.: Мир. 1978. ─ 119 с.
Теоретическое обоснование для подготовки к занятию
Аппара́т (ко́мплекс) Го́льджи ─ мембранная структура эукариотической клетки, органелла, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году (Fabene P.F., Bentivoglio M., 1998).
Рис. 1. Схема Аппарата Гольджи (А). Строение Аппарата Гольджи (Б)
Примечание: аппарат Гольджи ─ окруженные мембранами полости (цистерны) и связанная с ними система пузырьков. Функции ─ накопление органических веществ; «упаковка» органических веществ; выведение органических веществ; образование лизосом.
Аппарат (комплекс) Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединенных трубками стопок.
В Комплексе Гольджи выделяют 3 отдела цистерн, окруженных мембранными пузырьками:
1. Цис-отдел (ближний к ядру).
3. Транс-отдел (самый отдаленный от ядра).
Эти отделы различаются между собой набором ферментов. В цис-отделе первую цистерну называют «цистерной спасения», так как с ее помощью рецепторы, поступающие из промежуточной эндоплазматической сети, возвращаются обратно. Фермент цис-отдела: фосфогликозидаза (присоединяет фосфат к углеводу ─ маннозе).
В медиальном отделе находится 2 фермента: манназидаза (отщепляет манназу) и N-ацетилглюкозаминтрансфераза (присоединяет определенные углеводы ─ гликозамины).
В транс-отделе ферменты: пептидаза (осуществляет протеолиз) и трансфераза (осуществляет переброс химических групп).
Тонкое строение аппарата Гольджи (АГ). В электронном микроскопе видно, что аппарат Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне (рис. 1, 2); в виде стопки расположены плоские мембранные мешки (цистерны), количество таких мешков в стопке обычно не превышает 5-10. Между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре мембраны могут быть сближены (25 нм), а на периферии иметь расширения ─ ампулы, ширина которых непостоянна.
Рис. 2. Схема строения диктиосомы (по Ченцову Ю.С., 2010)
Примечание: П ─ проксимальная (цис-) часть; Д ─ дистальная (транс-) часть; В ─ вакуоли; Ц ─ плоские мембранные цистерны; А ─ ампулярные расширения цистерн.
У некоторых одноклеточных их число может достигать 20 штук. Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок (рис. 15.5). Между ними располагается средний или промежуточный участок АГ. Во время деления клеток сетчатые формы АГ распадаются до диктиосом, которые пассивно
и случайно распределяются по дочерним клеткам. При росте клеток общее количество диктиосом увеличивается.
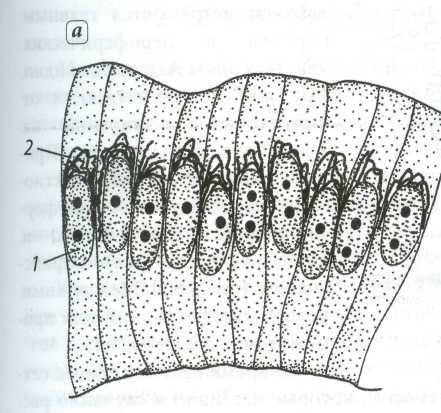
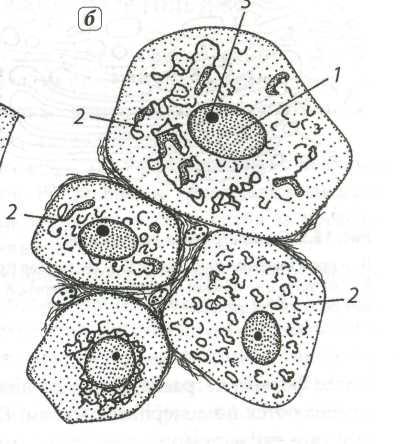
Рис. 3. Типы аппарата Гольджи (по Ченцову Ю.С., 2010)
АГ обычно поляризован в секретирующих клетках: его проксимальная часть обращена к цитоплазме и ядру, а дистальная ─ к поверхности клетки. В проксимальном участке к стопкам сближенных цистерн примыкает зона мелких гладких пузырьков и коротких мембранных цистерн.
Рис. 4. Аппарат Гольджи (АГ) в электронном микроскопе(по Ченцову Ю.С., 2010)
Рис. 5. Схематическое изображение компонентов аппарата Гольджи (по Ченцову Ю.С., 2010)
Примечание: 1 ─ ЭПР-АГ (ERGIC) ─ промежуточная зона; 2 ─ цис-зона, проксимальный участок; 3 ─ медиальный ─ средний участок; 4 ─ транс-дистальный участок; 5 ─ транс-сеть АГ.
В средней части диктиосомы периферия каждой цистерны также сопровождается массой мелких вакуолей около 50 нм в диаметре.
В дистальном или транс-участке диктиосом к последней мембранной плоской цистерне примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей, часто имеющих фибриллярную опушѐнность по поверхности со стороны цитоплазмы ─ это опушѐнные или окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе (от др.-греч. πίνω ─ пью, впитываю и κύτος ─ вместилище, клетка ─ захват клеточной поверхностью жидкости с содержащимися в ней веществами; процесс поглощения и внутри клеточного разрушения макромолекул).
Это так называемая транс-сеть аппарата Гольджи (TGN), где происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей ─ это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
При изучении толстых срезов клеток в мегавольтный электронный микроскоп было найдено, что в клетках отдельные диктосомы могут быть связаны друг с другом системой вакуолей и цистерн. Так что образуется рыхлая трехмерная сеть, выявляемая в световом микроскопе. В случае диффузной формы АГ каждый отдельный его участок представлен диктиосомой. В клетках животных часто с зоной мембран аппарата Гольджи ассоциированы центриоли; между радиально отходящих от них пучков микротрубочек лежат группы стопок мембран и вакуолей, которые концентрически окружают клеточный центр. Эта связь, вероятно, отражает участие микротрубочек в движении вакуолей.
Функции аппарата Гольджи. Наряду с белками в аппарате Гольджи осуществляется транспорт мембранных липидов.
1. Разделение белков на 3 потока:
● Цис-отдел (ближний к ядру); лизосомальный ─ гликозилированные белки (с маннозой) поступают в цис-отдел комплекса Гольджи, некоторые из них фосфорилируются, образуется маркёр лизосомальных ферментов ─ манноза-6-фосфат. В дальнейшем эти фосфорилированные белки не будут подвергаться модификации, а попадут в лизосомы.
● Медиальный отдел; конститутивный экзоцитоз (конститутивная секреция). В этот поток включаются белки и липиды, которые становятся компонентами поверхностного аппарата клетки, в том числе гликокаликса, или же они могут входить в состав внеклеточного матрикса.
● Транс-отдел (самый отдаленный от ядра); индуцируемая секреция ─ сюда попадают белки, которые функционируют за пределами клетки, поверхностного аппарата клетки, во внутренней среде организма. Характерен для секреторных клеток.
2. Формирование слизистых секретов (секреторная функция аппарата Гольджи) ─ гликозамингликанов (мукополисахаридов).
●Мембранные элементы АГ участвуют в сегрегации и накоплении продуктов, синтезированных в ЭПР, участвуют в их химических перестройках, созревании (перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран), (рис. 6).
●В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов.
●Главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, АГ ─ источник клеточных лизосом.
●Участие АГ в процессах выведения секреторных продуктов было очень хорошо изучено на примере экзокринных клеток поджелудочной железы. Для этих клеток характерно наличие большого числа секреторных гранул (зимогеновых гранул), которые представляют собой мембранные пузырьки, заполненные белковым содержимым. В составе белков зимогеновых гранул входят разнообразные ферменты: протеазы, липазы, карбогидразы, нуклеазы.
При секреции содержимое этих зимогеновых гранул выбрасывается из клеток в просвет железы, а затем перетекает в полость кишечника. Так как основным продуктом, выводимым клетками поджелудочной железы, является белок, то исследовали последовательность включения радиоактивных амино-кислот в различные участки клетки (рис. 7).
Рис. 6. Схема связи ЭПР, аппарата Гольджи с образованием и выделением зимогена из ацинарных клеток поджелудочной железы (по Ченцову Ю.С., 2010)
Примечание: 1 ─ переходная зона между ЭПР и АГ; 2 ─ зона созревания секреторных гранул; 3 ─ отделившиеся от АГ зимогеновые гранулы; 4 ─ их выход (экзоцитоз) за пределы клетки.
Рис. 7. Последовательность обнаружения [1-4) метки от 3 Н-лизина при синтезе и выведении белкового секрета из клетки поджелудочной железы (по Ченцову Ю.С., 2010)
Примечание: К ─ кровеносный апилляр, Ц ─ цитоплазма клетки; П ─ просвет железы. Стрелки показывают пути миграции метки.
Для этого животным вводили меченную тритием аминокислоту ( 3 Н-лейцин) и с помощью электронно-микроскопической радиоавтографии следили во времени за локализацией метки. Оказалось, что АГ является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Также подробно процессы синтеза и выведения белков были изучены на других клетках (молочная железа, бокаловидные клетки кишечника, щитовидная железа и др.), и были исследованы морфологические особенности этого процесса. Синтезированный на рибосомах, экспортируемый белок отделяется и накапливается внутри цистерн ЭПР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭПР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы (ЭПР-АГ комплекс). В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ.
●В аппарате Гольджи происходит не просто «перекачка» продуктов из одной полости в другую, но и постепенно идет их «созревание», модификация белков, которая заканчивается «сортировкой» продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
3. Формирование углеводных компонентов гликокаликса ─ в основном, гликолипидов.
4. Сульфатирование углеводных и белковых компонентов гликопротеидов и гликолипидов. В аппарате Гольджи происходит также сульфатирование глюкозаминогликанов и некоторых белков.
5. Частичный протеолиз белков ─ иногда за счет этого неактивный белок переходит в активный (проинсулин превращается в инсулин).
6. Транспорт веществ из эндоплазматической сети. Аппарат Гольджи асимметричен ─ цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки ─ везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭПР), на мембранах которого и происходит синтез белков рибосомами. Перемещение белков из эндоплазматической сети (ЭПС) в аппарат Гольджи происходит неизбирательно, однако не полностью или неправильно свернутые белки остаются при этом в ЭПС. Возвращение белков из аппарата Гольджи в ЭПС требует наличия специфической сигнальной последовательности (лизин-аспарагин-глутамин-лейцин) и происходит благодаря связыванию этих белков с мембранными рецепторами в цис-Гольджи.
7. Модификация белков в аппарате Гольджи. В цистернах аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т. д. Созревающие белки последовательно перемещаются по цистернам в органеллы, в которых происходят их модификации ─ гликозилирование и фосфорилирование. При О-гликозилировании к белкам присоединяются сложные сахара через атом кислорода. При фосфорилировании происходит присоединение к белкам остатка ортофосфорной кислоты.
В цис-зону аппарата Гольджи синтезированные в ЭПР белки попадают после первичного гликозилирования и редукции там же нескольких сахаридных остатков. В конечном итоге все белки там имеют одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетил-глюкозамина, шести молекул маннозы (рис. 8). В цис-цистернах начинается вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате олигосахариды на гидролитических ферментах, предназначенных для лизосом (олигосахариды, богатые маннозой), фосфорилируются, а олигосахариды других белков, направляемых в секреторные гранулы, или к плазматической мембране, подвергаются сложным превращениям, теряя ряд сахаров и присоединяя галактозу, N-ацетилглюкозамин и сиаловые кислоты. При этом возникает специальный комплекс олигосахаридов. Такие превращения олигосахаридов осуществляются с помощью ферментов ─ гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи.
По мере модификации белков в цистернах аппарата Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в трансучастке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз. Экзоцитоз (от греч. Έξω ─ внешний и κύτος ─ клетка) ─ у эукариот клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки) сливаются с внешней клеточной мембраной. При экзоцитозе содержимое секреторных везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с клеточной мембраной. Практически все макромолекулярные соединения (белки, пептидные гормоны и др.) выделяются из клетки этим способом.
Такое описание событий является только общей схемой участия аппарата Гольджи в секреторных процессах. Дело усложняется тем, что одна и та же клетка может участвовать в синтезе многих выделяемых белков, может их друг от друга изолировать и направлять к клеточной поверхности или же в состав лизосом.
Каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека («этажа» в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов.
Рис. 8. Пути вторичного гликозилирования белков в аппарате Гольджи (по Ченцову Ю.С., 2010)
Примечание: а ─ первичное гликозилирование в ЭПР; б ─ результат вторичного гликозилирования в АГ белков плазматической мембраны и секреторных вакуолей; в ─ результат вторичного гликозилирования белков лизосом; Ман ─ манноза; Асп ─ аспарагин; Гл ─ глюкоза; СК ─ сиаловая кислота; Га ─ N-ацетилглюкозамин; Гал ─ галактоза.
В цис-участке происходит фосфорилирование манноз в лизосомных ферментах и образуется особая маннозо-6-группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.
В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N-ацетилглюкозамина.
В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты (рис. 9).
Рис. 9. Локализация ферментов модификации белков в аппарате Гольджи (по Ченцову Ю.С., 2010)
Примечание: 1 ─ синтез белка в ЭПР; 2 ─ фосфорилирование лизосомных олигосахаридов; 3 ─ отщепление маннозы; ─ 4присоединение N-ацетилглюкозамина; 5 ─ присоединение маннозы; 6 ─ присоединение сиаловой кислоты; 7 ─ сортировка белков на рецепторах в транссети; 8 ─ лизосома; 9 ─ секреторная вакуоль; 10 ─ плазмалемма
8. Взаимосвязь аппарата Гольджи и эндоплазматического ретикулума. В разных областях аппарата Гольджи имеет место посттрансляционная модификация белков (рис. 9). Созревшие (модифицированные) белки переносятся везикулами в различные отделы клетки, такие, как лизосомы (4), цитоплазматическая мембрана (6) или секреторные пузырьки (5). Последние высвобождают свое содержимое к межклеточное пространство, сливаясь с плазматической мембраной (экзоцитоз). Эти транспортные процессы могут быть конститутивными, т.е. проходить постоянно, или регуляторными, т.е. управляться химическими сигналами. Направленность процесса в первую очередь зависит от сигнальной последовательности синтезируемого белка.
В образцах препаративно выделенных зон АГ при негативном контрастировании видно, что к проксимальной части диктиосомы примыкает сетевидная или губкообразная система мембранных полостей. Считается, что эта система (ЭПР ─ АГ) может представлять собой зону перехода элементов ЭПР в зону аппарата Гольджи.
Рис. 9. Аппарат Гольджи и шероховатый эндоплазматический ретикулум
Эндоплазматический ретикулум (ЭПР) ─ протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. В области ядра ЭПР сообщается с внешней ядерной мембраной. Между шероховатым и гладким ЭПР имеется морфологическое различие: мембраны шероховатого ЭР усеяны множеством рибосом, в то время как гладкий ЭР не имеет связанных рибосом.
Шероховатый эндоплазматический ретикулум (А) ─ место активного биосинтеза белков. Именно здесь синтезируются белки, которые будут функционировать в составе мембран, лизосом или секретироваться из клетки. Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭПР. Белки, синтезированные на шероховатом ЭПР (1), претерпевают посттрансляционные модификации (созревание белков). Они либо остаются внутри шероховатого ЭР в виде мембранных белков, либо транспортируются с помощью везикул (2) в аппарат Гольджи (3). Транспортные везикулы образуются почкованием мембран, а затем исчезают, сливаясь с ними.
Гладкий эндоплазматический ретикулум ─ эндоплазматическая сеть, не имеющая связанных рибосом (ГЭР), (рис. 10). Он занимает в клетке сравнительно небольшой объем.
Рис. 10. Аппарат Гольджи и гладкий эндоплазматический ретикулум
Выраженный ГЭР имеется в клетках с активным обменом липидов, таких, как гепатоциты и клетки Лейдига. Для ГЭР характерна замкнутая система разветвленных канальцев. ГЭР принимает участие в синтезе липидов. Биосинтез осуществляется ферментами, закрепленными на мембранах ГЭР. Здесь локализован синтез фосфолипидов и отдельные стадии синтеза холестерина. В ГЭР специализированных клеток эндокринной системы протекают различные стадии синтеза стероидных гормонов. В ГЭР локализованы также процессы метаболической трансформации ксенобиотиков. В этих реакциях принимает участие система цитохрома Р450, которую считают основной системой ГЭР. ГЭР выполняет функцию депо ионов Са 2+ , поддерживающего низкий уровень Са 2+ в цитоплазме. Эта функция более всего свойственна саркоплазматическому ретикулуму, специализированной форме ГЭР мышечных клеток (см. с. 326). В мембранах ГЭР локализованы управляемые Са 2+ -каналы и энергозависимые Са 2+ -насосы, а высокая концентрация ионов Са 2+ в цистернах поддерживается при участии Са 2+ -связывающих белков.
9. Сортировка белков в аппарате Гольджи. Через аппарат Гольджи проходит по крайней мере три потока синтезированных клеткой нецитозольных белков: поток гидролитических ферментов в компартмент лизосом, поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов, поток постоянно выделяемых секреторных белков. Существует специальный механизм пространственного разделения этих разных белков и их путей следования.
Аппарат Гольджи в нейронах. Спинномозговой узел котенка. Обработка четырехокисью осмия. Спинномозговые узлы млекопитающих находятся вблизи спинного мозга и содержат чувствительные нервные клетки. При малом увеличении видны крупные нервные клетки ─ нейроны или нейроциты (1) ─ округлой формы, располагающиеся преимущественно в периферических отделах спинномозгового узла.
Рис. 1. Аппарат Гольджи в нейронах. Спинномозговой узел котенка. Обработка четырехокисью осмия
Нейроны ─ наиболее крупные шаровидные клетки с отростками. Однако отростки нейронов выявляются только при специальной гистологической обработке и поэтому на данном препарате не видны. Между скоплениями нейронов находятся нервные волокна (2) и соединительная ткань. Вокруг нейронов видны в виде темных точек ядра (3) клеток-сателлитов или мантийных глиоцитов. Нейроциты отличаются друг от друга размерами, что связано как с действительными различиями их величины, так и направлением плоскости, через которую прошел срез. Надо найти нейроны, в которых видны структурные компоненты, и изучить их при большом увеличении (Б). Вокруг ядра (1) на светлом фоне нейроплазмы (2) выделяется черная извилистая сеть (3). При восстановлении четырехокиси осмия металлический осмий откладывается черным осадком на поверхность структурных компонентов аппарата Гольджи, придавая ему вид клубка нитей или сети (отсюда старое название этой структуры ─ внутриклеточный сетчатый аппарат). Аппарат Гольджи состоит из изогнутых и анастомозирующих между собой нитей и перекладин. Иногда эта сеть вплотную прилегает к ядру и охватывает его, образуя как бы корзинку (4). В других нейронах (5) она располагается несколько отступая от ядра. Встречаются нейроциты, в цитоплазме которых аппарат Гольджи состоит из отдельных палочек, чешуек, фрагментов (6), не связанных между собой и разбросанных по всей нейроплазме. Строение и расположение в цитоплазме аппарата Гольджи связаны с различным функциональным состоянием нейронов. В нервных клетках (7), ядро которых не попало в срез, аппарат Гольджи занимает центральную часть клетки. В поверхностно срезанных нейроцитах (8) он срезан тангенциально; в этом случае он имеет вид сети, занимающей своими перекладинами всю нейроплазму. Изучение аппарата Гольджи на светооптическом уровне свидетельствует, что сетевидная форма этого образования характерна для большинства клеток тканей взрослого организма. Электронная микроскопия показала, что аппарат Гольджи представляет собой сложную мембранную систему, в состав которой входят три типа структур: цистерны, микропузырьки и вакуоли. Эти структурные особенности аппарата Гольджи послужили основанием для его названия ─ комплекс Гольджи или пластинчатый комплекс. Участок цитоплазмы, в котором располагается этот органоид, называют «зоной Гольджи». Анализ ультраструктурных и биохимических особенностей пластинчатого комплекса в различных клетках животных и растений показал, что структура его элементов в разных объектах сходна и он функционально связан с процессами, обеспечивающими жизнедеятельность клетки. Такие наблюдения дают основание для причисления комплекса Гольджи к органоидам общего значения. Этот органоид особенно сильно выражен в клетках с интенсивным синтезом веществ (нервные, яйцевые, железистые). Комплекс Гольджи весьма активный органоид, участвующий в синтезе веществ, обеспечивающих рост и замену отмирающих частей клетки, а также потребности всего организма. Последнее достигается путем выделения синтезируемого продукта за пределы клетки.
источник